Największy z wielkich – tytan z Patagonii
 Słowo „dinozaury” oznacza w kontekście naukowym coś zupełnie innego, niż w znaczeniu potocznym. Wróbel jest dinozaurem, a pterodaktyl nie. Gdyby jednak przyjrzeć się tradycyjnemu, powszechnemu znaczeniu tego słowa, przychodzą na myśl wielkie, dawne gady, powolne olbrzymy zasiedlające pradawny świat na długo przed zapanowaniem człowieka.
Słowo „dinozaury” oznacza w kontekście naukowym coś zupełnie innego, niż w znaczeniu potocznym. Wróbel jest dinozaurem, a pterodaktyl nie. Gdyby jednak przyjrzeć się tradycyjnemu, powszechnemu znaczeniu tego słowa, przychodzą na myśl wielkie, dawne gady, powolne olbrzymy zasiedlające pradawny świat na długo przed zapanowaniem człowieka.
Pierwszy opisany rodzaj (nieptasiego) dinozaura, megalozaur, otrzymał nazwę odwołującą się do znacznych rozmiarów właśnie. Kolejny, iguanodon, nazwany został tak, bo jego ząb przypominał ząb legwana (Iguana) – z tym szczegółem, że był kilkakrotnie większy. Dziś wiadomo, że większość dinozaurów nie była aż tak duża, zaliczanie ich do gadów jest problematyczne, bo obejmują wszystkie ptaki, a zresztą słowo „gady” nie ma dobrej definicji. Z powyższego wynika też, że dinozaury nie wyginęły. Prawdopodobnie nie były też powolne (nie ścigałbym się z tyranozaurem, nawet jeśli rzeczywiście był padlinożercą). Rekord wielkości i masy zwierząt lądowych cały czas należy jednak do dinozaurów.
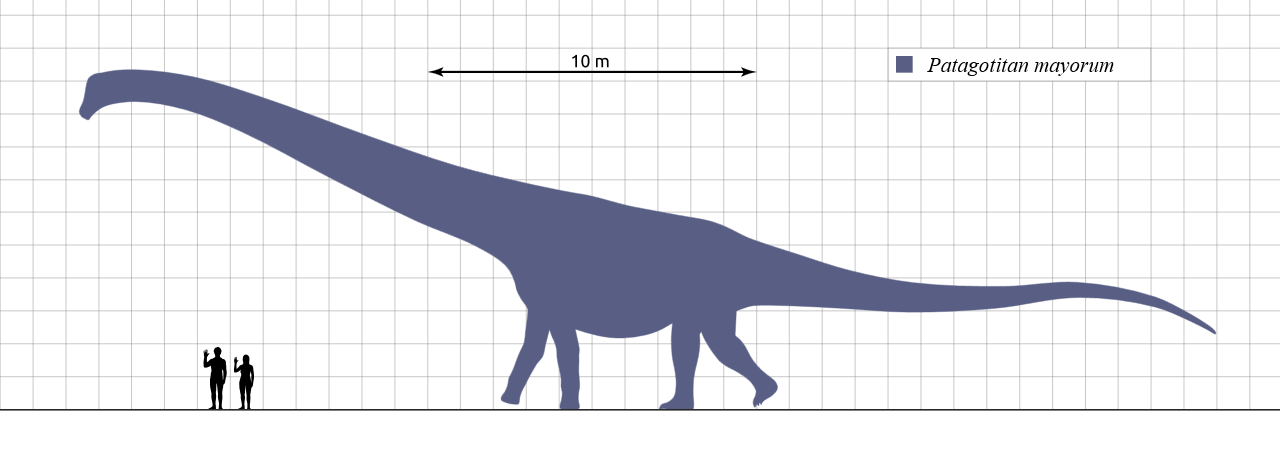
Obecny rekord pochodzi z Patagonii, argentyńskiej prowincji Chubut, gdzie w La Flecha znajdują się skały formacji geologicznej Cerro Barcino, powstałej pod sam koniec epoki kredy wczesnej, nieco powyżej 100 milionów lat temu. Ten wiek i zarazem piętro skał zwane są albem od nazwy rzeki we Francji. Tam pośród piaskowców mułowych i mułowców pozostałych po dawnej równinie zalewowej odkryto skamieniałe szczątki przynajmniej sześciu zwierząt. Okazało się, że były to młode, ale już dorosłe osobniki nieznanego jeszcze rodzaju zauropoda, które zapewne poniosły śmierć przy zbiorniku wodnym w różnym czasie.
Rodzaj zauropoda – to znaczy? Przejdźmy się krótko po systematyce. Wyobraźmy sobie kilkunastometrowego, dwunożnego tyranozaura o pysku pełnym ostrych zębów (najprawdopodobniej używał ich do miażdżenia kości padliny) oraz dziewięciometrowego, roślinożernego triceratopsa o trzech rogach i kryzie na szyi. Dinozaury dzielą się na dwie wielkie liniie: gadziomiedniczne (te bliższe tyranozaurowi niż triceratopsowi – może brzmi mało poważnie, ale tak się zwykle definiuje klady) i ptasiomiedniczne (odwrotnie, bliższe triceratopsowi). Żeby było śmieszniej, ptaki należą do gadziomiednicznych. Gadziomiedniczne dzielą się na zauropodomorfy (wyobraźmy sobie saltazaura albo inne podobne zwierzę, np. bardziej znanego diplodoka, o wielkim cielsku, długiej szyi zwieńczonej maleńką głową i długim ogonie) i teropody (tyranozaur, ptaszyska i to, co jest bliższe ptaszyskom niż zauropodomorfom).
Zauropodomorfy dzielimy na bardziej pierwotne prozauropody, jak plateozaur czy muszaur, z łatwością jeszcze wstające na dwie łapy i zwykle osiągające po kilka metrów długości. Ich szyja i ogon jeszcze nie są takie długie. Grupa ta żyła w późnym triasie i wczesnej jurze. Bardziej zaawansowane ewolucyjnie są zauropody, definiowane jako taksony bliższe saltazaurowi niż muszaurowi czy dzingszanozaurowi. Starsze z nich, jak wulkanodon, nie są jeszcze tak duże jak późniejsze. Ale już u jurajskich cetiozaurów zaznacza się charakterystyczna sylwetka, jeszcze wyraźniejsza u późniejszych diplodokokształtnych. Mała głowa umieszczona na długiej szyi, beczkowaty tułów, słupowate nogi i ogon, długi, niekiedy biczowaty (za wielkimi rozmiarami roślinożerców postępował wzrost drapieżników, od których trzeba się było czymś opędzać). Wszystkiego razem ze 30 metrów albo i więcej, niektóre szacunki mówią nawet i o 50 metrach w przypadku sejsmozaura czy amficeliasa.
Trochę inną sylwetkę miały Macronaria (dosłownie „duże nozdrza”), zwłaszcza Titanosauriformes. Nie były tak smukłe i górowały nad diplodokokształtnymi masą ciała. Przednie łapy były u nich często dłuższe od tylnych, jak u późnojurajskiego brachiozaura z Afryki, liczącego jakieś 23 metrów. O ile diplodokokształtne trzymały swe długie szyje zwykle poziomo (albo i nieznacznie opuszczone w dół), brachiozaury zadzierały swe głowy do góry na wysokość 12 metrów. Od jakichś bliskich krewnych brachiozaurów pochodzi jedyna w zasadzie grupa rozpowszechnionych w kredzie zauropodów, tytanozaurów. Zalicza się do nich kilka rodzin, paleontolodzy odchodzą już jednak od starego pomysłu Linneusza królestwo-typ-gromada-rząd-rodzina-rodzaj-gatunek. Jose Carballido i inni, autorzy opisu wspomnianego wyżej rodzaju z 2017 roku, podają jako jego klasyfikację pięć jednostek systematycznych, z których ani jedna nie należy do podstawowych według Linneusza.
Dinozaury mają rangę nadrzędu, zauropody – infrarzędu. Jak dalej przypisać rangi – nie mam pojęcia. Autorzy podają następnie tytanozaury, potem Eutitanosauria (czyli te „właściwe”, a więc bardziej zaawansowane ewolucyjnie tytanozaury), potem Lognkosauria (jakby nie można było wymyślać nazw, które da się wymówić). Nowy rodzaj ochrzcili Patagotitan mayorum. Nazwa rodzajowa oznacza „tytana z Patagonii”. Titanosauriformes często nazywa się przez odwołanie do pradawnych bóstw, olbrzymów z mitologii greckiej, pokonanych przez Zeusa i młodszych bogów (Paralititan, Gondwanatitan). Epitet gatunkowy wbrew pozorom nie odwołuje się do wielkości zwierzęta (maior to po łacinie większy, maiorum byłby dopełniaczem liczby mnogiej), lecz honoruje rodzinę Mayo, która udzieliła badaczom pomocy w czasie wykopalisk (jak widać, niekiedy warto wspierać naukowców).
Badacze spróbowali oszacować masę zwierzęcia. Nie można zrobić tego bezpośrednio (dostępne są tylko niektóre skamieniałe kości, a nalepek z prehistorycznej wagi dotychczas nie znaleziono). Stosuje się więc modele matematyczne, w których dużą rolę odgrywa często obwód kości ramieniowej bądź udowej, jest też metoda wolumetryczna (bazuje na objętościach). Na ile metody te są wiarygodne, mogą świadczyć oszacowania masy jednego z gatunków dikreozaura (wyjątkowo mały diplodokokształtny, sąsiad brachiozaura ze wschodnioafrykańskiej jury późnej) na 10-40 ton – jak na zauropoda to mało, dla porównania szacunek tą samą metodą Giraffatitan, uznawanego jeszcze niedawno za gatunek brachiozaura, to 74 tony.
Trzeba tu zwrócić uwagę, że przy tak dużych rozbieżnościach porównywać można jedynie wyniki uzyskane tą samą metodą. Wyniki dla Patagotitan wyniosły 69 ton i jest to najwyższy dotychczas uzyskany wynik tą metodą. Wcześniejszy rekordzista, dość blisko spokrewniony z nim Dreadnoughtus, był lżejszy o jakieś 10 ton, czyli 15 proc. (podkreślam – wedle pomiarów zastosowanych tą metodą, która dla wspominanego Giraffatitan dawała wyniki jakąś połowę mniejsze). Ogólnie maksymalna masa szacowana na podstawie kości ramieniowej i udowej dla tych dwóch rodzajów eutytanozaurów wynosi 74 tony. Wolumetria wskazuje na 45-77 ton. Ładny wynik. Dawniejszymi czasy spotykało się i szacunki znacznie mniejszego brachiozaura/Giraffatitan na 80 ton, ale – jak pisałem – ważna jest metoda, a stare szacunki rzadko się sprawdzają. No i uznanie ich oznaczałoby brak możliwości ogłoszenia rekordu.
Pozostaje jeszcze pytanie, czemu te zwierzęta były tak ogromniaste? Im jesteś większy – tym więcej drapieżników jest za małych, by cię zaatakować i pożreć. Jesteś wyższy – dosięgniesz wyższych liści. Łatwiej utrzymać stałą temperaturę ciała. No i dobór płciowy – samice wolą wyższych (a przynajmniej mniejszego konkurenta łatwiej odpędzić). To tylko luźno rzucane hipotezy. Faktem pozostaje trend ewolucji zauropodów w kierunku zwiększania rozmiarów ciała. Obserwowany też u wielu innych grup zwierząt, ale którzy z mieszkańców lądów, niekorzystający jak wieloryby z dobrodziejstw siły wyporu, mogą się z nimi równać?
Marcin Nowak
Ilustracja: A scale diagram showing the giant titanosaur Patagotitan mayorum, compared to some humans, autor Steveoc 86 na podstawie Carballido et al, 2017, Wikimedia Commons, na licencji CC BY-SA 4.0, zmieniono format pliku
Bibliografia:
Carballido Jose, Diego Pol, Alejandro Otero, Ignacio A. Cerda, Leonardo Salgado, Alberto C. Garrido, Jahandar Ramezani, Nestor R. Cuneo & Javier Marcelo Krause. A new giant titanosaur sheds light on body mass evolution among sauropod dinosaurs. „Proceedings of the Royal Society B”. 284, s. 20171219, 2017. Royal Society Publishing.
{kind=link}
Komentarze
Straciłem wątek. A gdyby tak coś z kryptografii?
Szanowny Panie Marcinie
Zwierzeta sa duze bo warunki im na to pozwalaja. Jednym z nich jest wyzasza niz obecnie koncentracja tlenu w atmosferze. Dawno temu bedac w museum dinozaurow zastanawialem sie nad wspolna dla nich i nas informacja genetyczna kodujaca na przyklad obecnosc 2 kosci podudzia czy jednej kosci udowej. Informacja genetyczna ktora dzielimy razem z dinozaurami jest przypomnieniem naszego wspolnego pochodzenia.
Slawomirski
Również jest to możliwe. Tlen pojawia się głównie w kontekście wyjaśniania istnienia olbrzymich karnońskich ważek i innych bezkręgowców, które nie mają płuc, a tylko mało wydajne tchawki.
Pluca niekoniecznie sa bardziej wydajne. Mechanizmy dostarczania tlenu dla mitochondrow sa rozmaite. Polarny gigantyzm dowodzi ze cisnieneie parcjalne tlenu rozpuszczonego w zimnej wodzie jest najistotniejsze. Antropocentryzm wymuszony religijnym wychowaniem szkodzi zrozumienu swiata.
Nie przypominam sobie, żeby Biblia coś na ten temat mówiła. Może ahabharata? Tym bardziej nie ma w tym antropocentryzmu, że najwydajniejsze są raczej płuca ptasie dzięki systemowi worków powietrznych. Natomiast owady mają tchawki, które uniemożliwiają i zwiekszenie rozmiarów ciała. A gigantym polarny wynika nie tylko ze stężenia tlenu w wodzie, jako że podobne zjawisko dotyczy zwierząt lądowych.
Szanowny Panie Marcinie
Najwydajniejsze pluca sprawiaja ze „wyzej ewolucyjnie” postawione foki staly sie pokarmem dla „niedorozwinietych” pozbawionych pluc rekinow. Przed chwila sam pan pisal o gigantyzmie owadow. Dlaczego tchawki maja limitowac zwiekszenie rozmiarow ciala. To srodowisko i ewolucja a nie tchawki sa odpowiedzialne za rozmiary ciala owadow. To samo dotyczy wszystkich zwierzat. Biblijny anrtopocentryzm nadal pana przesladuje.
Slawomirski
Foki są tak samo wysoko ewolucyjnie postawione jak rekiny.
Model przenoszenia tlenu i dwutlenku węgla w postaci gazowej rurkami (czyli tchawkami) do każdego zakamarka ciała, zamiast modelu przenoszenia go w postaci rozpuszczonej i przyczepionej do nośników typu hemoglobina sprawia, że im większe ciało, tym proporcjonalnie więcej tych rur musi być, aż do stanu, gdy rury te zajmują tyle miejsca, że na mięśnie czy inne tkanki już go brakuje.
Niby rozumowanie jest logiczne ale matka natura nie mysli logicznie tylko praktycznie i sprawila ze wazki mialy rozpietosc skrzydel do trzech stop a teraz sa takie jakie sa.
No właśnie sprawa wzięła się u stąd, że przy innej zawartości tlenu w atmosferze, te rurki mogą być proporcjonalnie węższe, czyli ostatecznie – obsługiwane przez nie ciało – większe.
Nie zawartosc tlenu ale jego cisnienie parcjalne i wynikajacy z tego gradient atmosferyczno-mitochondrialny dyktuja wielkosc organizmu. Prawa fizyki rzadza przeplywem gazow. Prawa biologiczne wymagaja optymalnego cisnienia parcjalnego tlenu w okolicy mitochondriow. Jedne i drugie musza byc respektowane przez ewolucje. To co pozornie nie jest mozliwe czasem w wyniku ewolucji staje sie faktem jak na przyklad oddychajace plucami ssaki zyjace w wodzie.
A czy w warunkach rzeczywistych ciśnienie parcjalne składnika mieszaniny nie zależy od zawartości tego składnika?
A płuca to bardzo stary wynalazek. I pojawił się nie tylko u ryb (niektórzy twierdzą, że to nie płuca powstały z pęcherza pławnego, ale odwrotnie) i ich potomków, ale też u ślimaków (także wodnych), a funkcjonalnie, to i niektórzy specjaliści od bezkręgowców twierdzą, że specjalne wyróżnianie odrębną nazwą płucotchawek jest przerostem formy nad treścią. Kluczowe jest to, czy dany worek ciała służy do wymiany gazowej z płynami ciała, które dalej obsługują tkanki, czy sam „ma ambicję” sięgnąć do wszystkich zakamarków tego ciała i dopiero tam dokonać tej wymiany.
Cisnienie parcjalne a nie FiO2 opisuje kwantytatywnie ilosc czasteczek tlenu. Pobieranie tlenu z atmosfery przez rogowke sprawia ze nie ma w niej naczyn krwionosnych. Rogowka oddych bez pomocy pluc .
Sprawa nie jest tak prosta. Nie ma gradientu powietrze-mitochondria. Po drodze jest kilka barier. Wpierw tlen musi przejść przez ścianę płuca do naczyń krwionośnych, tam znów przez błony do ertytrocytów. Stała wiązania Hb-O2 zależy od pH otoczenia, więc w miarę przepływu przez układ krwionośny to też się zmienia. Potem tlen musi zdysocjować od Hb, przejść przez kolejne błony, by w końcu dostać się mitochondriów, gdzie IV kompleks osadzi na nim elektrony i kationy wodoru. Wszystko to opisują logiczne prawa fizyki, wyrażalne przynajniej w teorii matematycznie (w rzeczywiście ukłąd jest skomplikowany i opisuje się je w sposób uproszczony).
Tam gdzie jest roznica cisnienia parcjalnego gazu istnieje gradient. Tlen nie jest wyjatkiem. Pomimo wiazania sie tlenu z Hb mierzymy we krwi cisnienie parcjalne tlenu. Tylko bardzo mala ilosc tlenu jest rozpuszczona fizycznie we krwi. Istnienie gradientu wymusza transfer tlenu z atmosfery do enzymow oddechowych mitochondriow gdzie molekularny tlen zmienia sie w wode. Mozna powiedziec ze znikanie molekularnego tlenu w mitochondrach sprawia ze jest on zasysany z atmosfery.
Nieprawda. Gradient ma sens w przypadku dwóch sąsiadujących ze sobą obszarów o różnym stężeniu/ciśnieniu. Tutaj mamy szereg gradientów tworzących złożony układ. Nie da się pominąć tego, co znajduje się pomiędzy matrix mitochondrialną a atmosferą.
Roznica cisnienia parcjalnego tlenu miedzy atmosfera a miejscem gdzie molekularny tlen zamienia sie w wode stanowi gradient. Dwa punkty pomiaru okreslaja gradient.
Oczywiscie miedzy tymi punktami pomiaru cisnienie parcjalne tlenu molekularnego moze byc i jest mierzone jak na przyklad w powietrzu pecherzykow plucnych czy we krwi tetniczej czy gdziekolwiek na drodze z atmosfery do pojedynczej komorki organizmu. Nie sadze aby mozliwy byl pomiar cisnienia parcjalnego tlenu w mitochondrialnym matrix. Przy zalozeniu ze caly molekularny tlen jest przerobiony przez enzymy oddechowe na wode cisniene parcjalne tlenu w tej czesci mitochondriona powinno byc zero. Czyli gradient atmosferyczno mitochondrialy jest taki sam jak cisnienie atmosferyczne tlenu.
Dziekuje za dyskusje.
Slawomirski