Nowotwór – atawizmem?
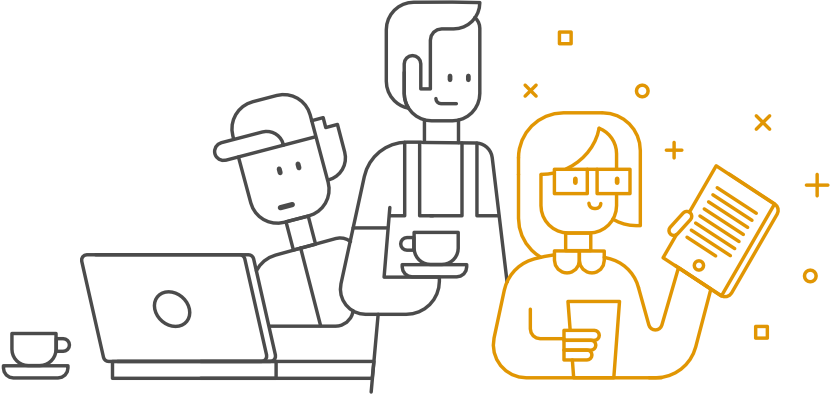
Termin „atawizm” w biologii oznacza pojawienie się u danego osobnika cech – zwykle anatomicznych lub zachowania – typowych dla jego odległych przodków, nieobecnych w populacji, do której należy. W przypadku człowieka najczęściej wskazuje się szczątkowy ogon czy żebra szyjne, poza tym palce konia czy kończyny tylne waleni. Rzadziej spotyka się pogląd, że przykładem atawizmu są… nowotwory.
Najszerzej znany model nowotworzenia bazuje na kolejnych mutacjach, poczynając od zachodzących w specyficznych grupach genów: związanych z regulacją cyklu komórkowego (a więc i kontrolą podziałów komórki), wykrywaniem i naprawą DNA, apoptozą (kontrolowana śmierć komórki). W niezbyt licznych przypadkach (np. retinoblastoma) pierwsza mutacja może być dziedziczona.
Mutacje z zasady zachodzą losowo i zazwyczaj nie mają wpływu na funkcjonowanie komórki. Zmiana genu kodującego barwnik oczu w komórce mięśnia sercowego nie wpłynie na EKG.
Jeśli jednak mutacja zajdzie w genach wspomnianych grup, komórka może wyzwolić się z kontroli i zacząć dzielić się nadmiernie. Może nie umrzeć, kiedy jest to wymagane dla dobra organizmu. Może dzielić się dalej, mimo że jej materiał genetyczny został uszkodzony. Upośledzenie naprawy DNA prowadzi do akumulacji mutacji. Komórki coraz mniej służą reszcie ciała, są coraz bardziej niezależne. Oczywiście w dużym uproszczeniu (którego nadmiar mi tu nie raz zarzucano, ale to chyba nie miejsce na rozwodzenie się nad mechanizmem działania białka p53 ani kinaz cyklinozależnych czy naprawy DNA).
Komórka się dzieli, w różnych jej potomkach zachodzą kolejne mutacje. Mamy populację coraz bardziej różniących się komórek w środowisku o ograniczonych zasobach – taką sytuację opisuje teoria ewolucji przez dobór. Z drugiej strony liczne nowotwory wykazują znaczne podobieństwo – wzrost w formie guza, nowe unaczynienie, niekiedy zdolność do przerzutowania czy oporność na chemioterapię. Z ewolucjonistycznego punktu widzenia opisalibyśmy to jako konwergencję – rozwój podobnych cech u oddalonych od siebie organizmów. Typowy przykład: opływowy kształt ciała wieloryba i ichtiozaura, podobieństwo wilka i wilka workowatego.
Niektórzy badacze widzą w podobieństwach łączących nowotwory o różnym pochodzeniu coś więcej. Szybki podział komórki, odporność na toksyny, zdolność do wzrostu w środowisku o niskim pH, szybkie tempo mutacji – cechy te występują także u mikroorganizmów. Wszyscy pochodzimy od mikroorganizmów, jakichś prekambryjskich eukariontów, które zaczęły tworzyć coraz bardziej skomplikowane kolonie, z których powstały organizmy wielokomórkowe. Kiedy dany gen przestaje być użyteczny, nie znika w magiczny sposób z genomu. Często jest w jakiś sposób nieaktywowany (np. przez produkty innych genów czy mutację uniemożliwiającą mu działanie). Oczywiście metodą prób i błędów inaktywowane są także nader użyteczne geny, ale wtedy noszący taką mutację organizm ma mniejsze szanse przekazania jej następnemu pokoleniu (a niekiedy nawet przeżycia). Gdyby przypadkowo zostały one ponownie aktywowane, będziemy mieli do czynienia z atawizmem.
A więc geny związane z przeżyciem mikroorganizmu w prekambryjskim oceanie mają się w tym ujęciu aktywować i prowadzić do zachowania się komórki, uprzednio trybiku w wielkim organizmie, ponownie jako niezależny organizm. Który dzieli się, kiedy ma taką możliwość, a nie kiedy cząsteczka sygnałowa połączy się z jej receptorem, udzielając jej zgody na podział. Nowo spłodzone potomstwo mikroorganizmu, takiego, z którego na drodze stopniowych przemian ewolucyjnych powstać miały ja, słoń i sekwoja olbrzymia, miałoby pewnie tendencję do życia w kolonii. Jej odpowiednikiem w przypadku nowotworu ma być guz. Zdolność przeżycia w zakwaszonym oceanie stanowić ma o przeżywalności w kwasicy wywołanej intensywnym metabolizmem.
Badacze wskazują na jeszcze jedną ciekawą właściwość mikroorganizmów. Otóż powoływanie do życia swoich dokładnych kopii opłaca się najbardziej w stabilnych warunkach. Mszyce w lecie rozmnażają się przez partenogenezę (rozwój kopii matek z niezapłodnionych jaj). Jednak jesienią samce okazują się przydatne – a nawet niezbędne do wydania zróżnicowanego genetycznie potomstwa. W zmiennych warunkach jesieni zwiększa to szansę, że część potomstwa okaże się lepiej przystosowana i przeżyje. Podobnie mikroorganizmy zwiększać mają częstość mutacji w trudnych warunkach. Zjawisko to odpowiadać ma niestabilności materiału genetycznego komórek nowotworowych.
Nie ma obecnie badań pozwalających jednoznacznie stwierdzić, który z modeli (przypadkowych mutacji, doboru i konwergencji versus atawizmu) jest właściwy. Być może po części oba, gdyż się nie wykluczają. A więc po trochu losowe mutacje, po trochu włączenie się nieużywanych od pokoleń genów. Rozstrzygnięcie problemu można by zapewne wykorzystać w tworzeniu i przewidywaniu wyników leczenia. Inaczej walczy się z całkowicie nieprzewidywalnym, działającym losowo przeciwnikiem, inaczej z realizującym dawno zapomnianą strategię.
Marcin Nowak
Bibliografia
Thomas F, Ujvari B, Renaud F, Vincent M: Cancer Adaptations: Atavism, De Novo Selection, or Something in Between? Bioessays 2017 Aug;39(8). doi: 10.1002/bies.201700039. Epub 2017 Jul 10
H%26E_magn_200x.jpg?uselang=pl){kind=link}
Komentarze
@MNowak
Rozstrzygnięcie problemu można by zapewne wykorzystać w tworzeniu i przewidywaniu wyników leczenia. Inaczej walczy się z całkowicie nieprzewidywalnym, działającym losowo przeciwnikiem, inaczej z realizującym dawno zapomnianą strategię.
Na przyklad?
Na cos trzeba umrzec. Zycie wieczne jest absurdem. Kontekst jest wazny. Dzieci i mlodzi ludzie nie powinni umierac na nowotwory. Przed nimi cale zycie. Leczenie tych nowotworow jest bardzo potrzebne. Na ten cel warto wydawac pieniadze.
Przykladow szanowni autorzy tej dziwacznej koncepcji nie podają. Jednakże gdyby okazała się zasadna, można by przenieść wyniki badań z mikroorganizmów.
@MNowak
Wpis jest w calosci streszczeniem artykulu podanego w bibliografii lacznie z opiniami? Nie bylo to jasne z tekstu.
@Marcin Nowak
to chyba nie miejsce na rozwodzenie się nad mechanizmem działania (…) kinaz cyklinozależnych
Czemu nie? Opisanie mechanizmu działania kinaz cyklinozależnych jest pewnie tylko o szczebel trudniejsze od mechanizmu działania parkietów cyklinozależnych. W każdym razie może czytelnicy podejmą ten trud i coś (połowicznie?) ogarną.
Mnie od dawna fascynuje to, że nowotwory są chyba nieprzypadkowe i np. hist-pat pozwala odróżnić poszczególne inne (morfologicznie) rodzaje komórek nowotworowych w tej samej okolicy cielska.
Pacholęciem na studiach będąc słyszałem, że gen(y?) odpowiedzialny/e za powstawanie polipów w ludzkim jelicie przednim lub tylnym (a więc ektodermalnym – anatomom antropocentrycznym przypominam, że w ogólnej anatomii zwierząt jelito przednie to ten kawałek, który u małej grupki zwierząt określany jest jako jama nosowa, jama ustna itp.) są ewolucyjnie te same, co geny odpowiedzialne za powstawanie polipów potomnych u parzydełkowców (osobom kończącym znajomość taksonomii mniej więcej na maturze z moich czasów – to grupa jamochłonów). Innymi słowy, polipy, a więc często początek raków (bądź gruczolaków), to atawizm pączkowania.
@karat
Tekst jest przedstawieniem poglądu głoszonego przez autorów artykułu, z pewnymi dodatkami ode mnie, żeby tekst był zrozumiały i stanowił pewną całość
@Gammon No.82
31 MAJA 2020
Nie wiem, czy bym się poważył… Na studiach ćwiczeniach z genetyki własnie na genetyce nowotworów wiekszość osób poległa i z mojej ponaddziesięcioosobowej grupy do zaliczenia ćwiczeń dopuszczono 4
@MNowak 12:04
To powinno byc wyraznie zaznaczone w tekscie, nie wystarczy luzne dodanie bibliografii.
Tak sie zastanawiam, kładę palce wskazujące na klawiaturze i …zabieram.
Ale, co miałam stracić to straciłam a więc…
Jestem jak zeszyt ćwiczeń dla R(r)ezydentów, jestem właścicielka
dwóch Bliznowców, jeden jest już nawet po 2o tce i operacyjny,
ma 12 cm blizna, guzki na niej są wypukłe i wklęsłe rozlewające się, drugi maleńki
na zeszłorocznej bliźnie.
Ukrywam je i przed sobą też, bezwolnie zasłaniam brzuch, wzbudzam tym ciekawość
przechodniów, zdarza sie że pytają czy mnie coś boli, przed współpracownikami
zasłaniam go koszulka bokserka, by mnie nie napiętnowały, wiadomo jakie są kobiety…
Każde z mojego rodzeństwa miało już kontakt ze skalpelem, a takie blizny mam tylko ja.
W „Stranger Things” w galerii Nastka wyzbyła sie intruza z łydki swej, na co Mike
powiedział jej że” z bliznami jest jeszcze ładniejsza”….
Czy bliznowce dziedziczymy?!
Nie warto marnowac czasu na pytanie autora czy nowotwor jest atawizmem. Nawet gdyby byl atawizmem to nie zmieni to faktu ze nowotwory czesto prowadza do smierci czy permanentnego uszkodzenia. Czasem lepiej jest milczec niz spekulowac w konfrontacji z ludzka tragedia. Pisac warto gdy ma sie cos istotnego do zakomunikowania. W przeciwnym razie slowa zamieniaja sie w niezroruzumialy/zarozumialy tekst w kontekscie nowotworowego nieszczescia. Czy rodzice piecioletniego dziecka umierajacego na raka lub mloda matka dwojki dzieci z terminalnym rakiem piersi znajda w tym tekscie pocieche?
@Slawomirski
A. Pytanie nie jest autora lecz badaczy ktorych nazwiska figuruja w bibliografii, a korzy napisali artykul stanowiacy przeglad zagadnienia.
B. To artykul opublikowany w czasopismie naukowym, w ktorym publikacje nie maja intencji pokrzepienia serc szczerze cierpiacych, sa adresowane do osob zajmujacych sie tematem: zawodowo, nie personalnie i nie emocjonalnie. Bez istnienia takich ludzi cierpiacy beda cierpiec dalej.
C. Samo zagadnienie jest w miare ciekawe: przewaznie mysli sie o nowotworach jako o wyniku przypadkowych mutacji, w wyniku ktorych komorka przestaje byc podobna do swojej zdrowej formy oraz, co bardzo wazne, przestaje reagowac na rozne systemy kontrolne tak wewnatrz samej komorki, jak i poza nia. W tym artykule przedstawiona jest inna mozliwosc, ze to uwolnienie sie spod kontroli, jakby anty-socjalizacja komorki nowotworowej, dotyczy roznych starych genow stlumionych od bardzo wielu pokolen w toku ewolucji. Jesli tak mozna by sie np zaczac zastanawiac czy badania w tym kierunku moga doprowadzic do zrozumienia jakiegos aspektu chorob nowotworowych lub przynajmniej niektorych z nich, poniewaz w takiej sytuacji nowotwory beda moze mniej przypadkowe. Co prawda nie jestem pewna czy wynikiem akurat tego rozwazania bedzie jakis insight, tym bardziej ze, wg streszczenia, autorzy siegaja od razu az po bakterie, przeskakujac lekkim krokiem prawie cala ewolucje, co odbiera pojeciu atawizu wiekszosc sensu, ale mniejsza z tym 😉
Przyklad @ppanka o polipach szalenie ciekawy, ale nic na ten temat nie znajduje, niestety.
W zakladce „O AUTORACH” (prawa gorna) figuruje mail redaktora odpowiedzialnego za ten blog. Proponuje pisac do niego z prosba o poprawki szaty graficznej, dodajac ze w odroznieniu od innych blogow, tu autorzy nie podjeli sie przekazac naszych prosb i dlatego musimy indywidualnie 😉 Im wiecej osob napisze tym lepiej..
„Atawizm (łac. atavus – przodek) – regresja ewolucyjna, pojawienie się u danego organizmu nietypowych dla niego cech bądź zachowań, charakterystycznych dla jego odległych przodków.” (Wikipedia)
@Karat
„…dotyczy roznych starych genow stlumionych od bardzo wielu pokolen w toku ewolucji.”
Przepraszam ze tak pocięłam, ale atawizm nasunął mi bliznowca
bo jest łagodna odmiana nowotworu, z założenia dotyka ciemniejsze pigmenty
skóry, ja powiedzmy cieszę sie barwą skóry typowo europejską…
A mój przodek mógł być np. Algierczykiem, Libijczykiem, Egipcjaninem…
„W Tanganice rządzonej przez Niemców niewolnictwo zniesiono dopiero 1922 roku.”
https://historia.uwazamrze.pl/artykul/1147605/krotka-historia-niewolnictwa
Skądś przyszedł, może i go przywlekli…a pamiętam z ulic Paryża, z jakiś zdjęć, czy książek
ciała ciemnoskórych pokrytych bruzdami.
Skoro potrzebuję odpowiedzi to taka mnie zadowala, i mam nadzieję ze nie dotknie juz moich dzieci czy wnuczki…
A te polipy to przecież miało być coś błahego?!
@R.S.
Zboczę na moment z tematu, ale muszę…
R.S. jeśli jeszcze dłubiesz się z tą elewacją mam pomysł na nią?!:
https://www.youtube.com/watch?v=IIzemxXzIQc
Coś pięknego, a nuta nadaje się do porannej kawy we dwoje…
Co to jest „kliniczny punkt widzenia”
@karat.
1 CZERWCA 2020
20:24
Ladnie to tlumaczenie wyszlo. Niektorzy potrafia nas przekonywac do niczego. Maja takie zdolnosci sa bardzo inteligentni z aparycja i gladka jak lody waniliowe osobowoscia. Na Dzikim Zachodzie w czasch goraczki zlota sprzedawali olej z weza na wszystkie mozliwe dolegliwosci. Ci ludzie sa z nami od zawsze. Robia kariery naukowe bez robienia nauki. To co robia jest obrabianiem nauki na potrzeby ich kariery. Aby uniknac zaplatania w sidla tych osob watro pamietac o podziale nauki na science i bs science(bad science). Wartosciowych odkryc naukowych jest niewiele sa one jak perelki na dnie oceanu bs science.
@Slawomirski
Prawda, ale tez:
1. Watpie czy jest droga na skroty – musi byc duzo malenkich kroczkow po to by mogl zaistniec ten jeden wiekszy, „perelka”.
2. Wartosciowe odkrycie niekoniecznie jest odkryciem majacym praktyczne zastosowanie.
@karat.
2 CZERWCA 2020 9:44 punkt 2.
<3 bo co tu długo mówić
@Gammonie
Czyżbyś czekał 12 godzin i 2 minuty, by 45 razy dotknąć / musnąć klawiszy / u…?
Droga Bohatyrowiczowo,
Dzieki za pamiec I podeslana kombinacje malarsko muzyczna.
Byc moze Cie rozczaruja, ale nic nie zastapi swiergotu ptakow I widoku kwitnacych azalii, rododendronow, peoni drzewiastych, magnolii wraz z kakafonia ich zapachow. Szczegolnie teraz gdy ani dzwiekow I zapachow nie zatruwa oddalona o mile autostrada.
Najmocniej przepraszam ludzi mieszkajacych w miastach bowiem Covid jest tam duzo bardziej przerazajacy I uciazliwy niz u mnie na pseudo-wsi, wiec prosze aby nie traktowac wpisu jako a mnie to wszystko co sie dziej guzik obchodzi. Wrecz przeciwnie, obchodzi I to bardzo.
Projekt to nie fasada a wrecz caly dach (finska blacha dachowka), ktory zostal powyginany w wyniku oberwanej trzystu kilogramowej galleezi. Przy okazji wymiany odkrywam cala ekosfere, ktora sobie zyla pod dachem, milo I bezpiecznie. Wiewiorki drzewne, weezze, termity, pszczolly I inne. Ta litera ‘I’ to wynik filtru Microsoft Worda, ktory namolnie przerzuca ‘i’ na duza literee.
O frustracjach naukowych nie bardzo chce mi sie wspominac, bowiem u nas pllonaa miasta I gina ludzie.
Probuje zaaplikowac sobie odwyk blogowo internetowy I z gory przepraszam za latencje odpowiedzi.
@R.S.
Alezz bardzo proszee i calla przyjemnosss po mojej stronie…
Mam ochotee i to zrobiee porooownam Covid do clipu Katie Melua
– The Flood
https://www.youtube.com/watch?v=4E4-9yKTv_I
Inpiracjaa jest nowa arka – pociaag z Netflixa
I tak, nie siejaac zgorszenia, mamy tam do wyboru:
skały, pałka w dlloniach imituje samobronee, walkee niczym wiosłem przeciwko falom, mamy i arkee przypominajaacaa trumnee gorszego sortu, konnnczymy
dryfujaacym fortepianem, jej narzeedziem pracy, moim marzeniem…
Kaleczymy siee, toniemy, opadamy samotnie na dno, prooobujemy utrzymaccc
siee w siodle, nie daccc odebraccc sobie marzennnn….
Przy Katie sllowach „Blame, no one is to blame ” popuszcza mi spina pod llopatkami, podejmujee walkee o swe lepsze zzycie, jego jakosscc przedewszystkim!!!
Pozdrawiam. 🙂
@R.S.
Pisząc powyższy tekst skupiłam sie na rozbijaniu, umknęła mi przy tym ilość
literówek, i nierozbitych liter, choć i treść kłębiła mi się inna w myślach
światło dzienne ujrzała tylko część, samoobrona miała być naszym układem
odpornościowym, kaleczenie przy roztrzaskiwaniu, niemocą się niczego chwycić, zaciągnąć itd
Popracuje nad Twoim sposobem, by chodziło po twojemu ale tak jak ja chce a nie chwila nieuwagi we mnie…
Gammon
Ciesze sie ze jestesmy zgodni 🙂
Droga Bohatyrowiczowo,
Millo mi zze proobujesz. Wnioskujee, zee nie lubisz chodzicc utartymi ssciezzkami , tylko takimi ktoore Ci odpowiadajaa.
W praktyce po zainstalowaniu odpowiedniej wklladki w softwarze (kernel), zmiana na literee z akcentem beedzie zachodzilla automatycznie w wyniku ktoorej rezultat, wyglad sllowa jest identyczny.
Calla koncepcja opiera siee na enkodowaniu liter z grupy Latin-2 za pomocaa klasycznej klawiatury QUERTY przy jak najmniejszym wysillku.
Dawno temu przeprowadzono pomiary podczas ktoorych zmierzono, zze reeka czllowieka piszaacego na klawiaturze QUERTY przez osiem godzin pokonuje dystans okollo osiemnastu mil.
Zatkallo mnie!
Problem zmeeczenia proobowano rozwiazacc stosujaac klawiaturee w ukladzie Dvoraka.
W poroownywalnym tesscie piszaac na klawiaturze Dvoraka ruch reeki zostall zredukowany do jednej mili. Czyli fatyga nadgarstka zmiejszylla sie azz osiemnasscie razy.
https://en.wikipedia.org/wiki/Dvorak_keyboard_layout
Klawiatura jednak siee nie przyjella.
Sposob enkodowania postanowilem nazwacc “DOUBLE KEYSTROKE ENCODING OF POLISH CHARCTERS”.
Mozze powinienem zarejestrowacc sobie kredyt jako wynalazca?
Microsoft lubi podkradacc pomyslly i ogllaszacc SSwiatu jako swoje.
Wiem coss o tymz wllasnego dosswiadczenia 🙁
A o tym jak elegancko w sytemie operacyjnym wprowadzicc opcjee DOUBLE KEYSTROKE, trzeba by posswieecicc cally wsteepniak i zapoznacc czytelnikoow z magiaa programowania systemowego.
Programowanie systemowe to nie to samo co pisanie aplikacji, lub macros.
Uwierz mi, zze to naprawdee wyzzsza szkolla jazdy.
Wssrood populacji programistoow softwaru stosunek systemowcoow do reszty jest jak 1:5000.
Na zakonnczenie podam link dajaacy przedsmak monitorowania wprowadzonych znakoow podczas pisania na klawiaturze :).
https://www.codeproject.com/Articles/43144/Keystroke-Monitoring
Wybacz ale wyllaczee siee z dalszej dyskusji, jeszcze raz dzieeki za uwagee.
@R.S.
Myssslee zze dallam Ci kapkee odetchnaaccc, rozumiem meeskaa
potrzebee izolacji…
Sssciaagnellam sobie ten program na lapka ale mooj Windows 10,
go nie wesprze zdaniem mojego Syna, ucznia technikum informatycznego,
kojarzy zze Wielki Brat czyt. wychowawca praktyczny mooowill o tym programie
ale to dopiero w trzeciej klasie beedaa mieccc go na tapecie…
Moooj laptop dogorywa wiec nie beedziemy ruszaccc oprogramowania,
beedee o tym pamieetaccc przy kupnie nowego…a nuzz siee przyda.
I tak za Twoim posssrednictwem pobrodzillam po ssswiecie syna…dzieekujee.
Ja nie chce wyprowadzicc gloownego tematu wsteepniaka nad ktoorym natrudzill sie kolega Nowak na boczny tor informatyczny. Staad moja wstrzemiezzzliwosscc.
Bardzo, krootko dla przypomnienia.
Podwoojne litery (double keystroke encoding) , to propozycja (by R.S.), metody enkodowania polskich znakoow diakrytycznych. Mozzna posluzzycc siee analogiaa funkcjonalnosc click vs. double click w myszy. System operacyjny windows rozroznia te dwie metody clickania myszaa.
W przypadku klawiatury takiego dynamicznego rozroznienia nie wprowadzono, staad potrzeba podllaczenia dodatkowego filtru (kernel), analizujacego szybkosc uderzenia w klawisze liter a, e, c, o, n, s, z oraz l i dokonania analizy.
Przyklladowo po zainstalowania drivera filtru klawiatury, dwa szybkie uderzenia tej samej litery ‘l’ , ‘l’ w odsteepie np. mniejszym niz 0.1 sekundy powinno dacc ‘ł’ , natomiast jessli interwall bedzie dluzzszy nizz dziesiaata czeesscc sekundy dostaniemy dwie litery ‚ l’ jedna po drugiej. Czyli mozzemy bardzo prosto zakodowacc sllowo troł lub troll bez wciskania Alt-X, Ctrl czy innego wygibasa.
Syn ma racjee, przytoczony link pokazuje technologiee drivera filtrujaacego klawiature w oparciu o stary model WDM ( Windows Driver Model) – ktoory byl stosowany w systemach operacyjnych Windows 98 – 2000. Windows 10 jest oparty o nowszaa technologie driveroow WDF.
Sugerujee trochee lepszy tutorial pokazujacy filtr klawiatury jest w linkach ponizzej.
Wielki brat mozze oprzecc siee na tych dwooch przyklladach.
https://www.youtube.com/watch?v=JVWtFsmOkA0
https://www.youtube.com/watch?v=UlzctiY2NSg
Mimo, zze tutoriale majaa narracjee jak flaki z olejem, zdolnaa usspicc kazzdego po minucie oglaadania, merytorycznie saa bardzo dobre!
Uwaga fonetyczna IRP wymawiamy tak jak po polsku a nie jak komentator AJ-AR-PI
Tutoriale to wprowadzenie do tematu filtrowania.
Kodu do double keystroke encoding tam nie znajdziesz. Ten kod trzeba napisac.
Mam nadziejee zze teraz to naprawdee zaimponujesz synowi 🙂 i Wielkiemu Bratu
Powodzenia.
Żałuje że poruszyłem ten temat – który RS doprowadził do absurdu – sorry.
@observer
Może ten klasyk, który mi się nieodparcie nasuwa w takich sytuacjach, cię jakoś pocieszy:
https://w-zaciszu-biblioteki.blogspot.com/2012/07/jak-zapamietac-duze-liczby-czyli.html
Zastanawiam się jak zapamiętać PIN do nowej karty SIM – 7842. Chyba prędzej zmienię na stary, który pamiętam od lat 🙁
Wręcz przeciwnie Markocie,
Osobiście jestem wdzięczny.
Bez Twojego kodowania ‘ś’, nie zwróciłbym uwagi a tym bardziej nie próbował rozwiązać problemu.
A ortograficzne wyjątki były dość pomocne.
@markot
Uwielbiam humor Szwejka, co kilka lat czytam go ponownie.
Ja już pamiętam Twój PIN, przed podjęciem zadzwoń do mnie.
@observer
Dzięki! 😀 😀 😀
PS
Postanowiłem zmienić na 4268. Albo to było 4286?…
Znowu mam kalendarz z boku kolumny. Tumiwisizm i bylejactwo wśród IT.